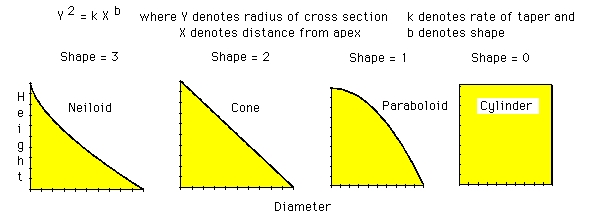
Taper of a solid body of regular outline is governed by the values of 'rate' and 'shape' in the expression:
diameter^2 = rateheight^shape
where shape determines the way the body tapers (i.e. form or shape) and rate the rate of taper within that form.
If we knew more about the factors determining stem form, we might be in a better position to control it: a cylindrical stem would be an obvious advantage in utilisation! Certainly species, genotype, age, competition, site (especially wind exposure), silvicultural treatment and size and structure of the crown are all important. Of these factors, the crown, particularly crown length, plays a decisive role in determining stem form.
Some general patterns are:
| Tree situation | Ring area growth | Ring width |
| Free growing trees, predominants or vigorous trees growing in gaps Crowns strongly developed | Increases basipetally | Increases basipetally |
| Stand grown trees not overtopped but side growth of crown hindered e.g. codominants | Approx. equal growth along length of stem | Decreases basipetally |
| Overtopped stand grown trees, e.g intermediate and suppressed trees | Decreases basipetally | Decreases basipetally |
Although crown size and development definitely determine stem growth, many variables (age, site, location in the stand, etc.) intercede to further determine how this growth is distributed over the stem.
A pronounced region of minimum ring area growth is often encountered on the branch free bole. In young stems, it occurs near the ground shifting upwards towards the crown base in dry years and downwards towards the stem base in wet years, i.e. it varies with the prevailing growth conditions and parallels the region of maximum area growth near the crown base.
Regardless of the ring width pattern along the stem, a relatively high degree of taper is always found within the crown. This is because of the progressive increase in branch numbers downward from the apex and the cumulative contribution of these branches to stem growth.
Cultural measures such as thinning and pruning can markedly alter the incremental growth pattern of individual trees by artificially creating new growth conditions. Thinning converts a stand-grown tree to a simulated open-grown condition, and growth will shift downward on the stem in response to the increased crown size and exposure. Pruning, on the other hand, converts an open-grown or large-crowned tree to a simulated stand-grown condition, and growth will shift upward on the stem in accordance with the new crown position. Again, these responses are not invariable and a change in form often fails to occur following a silvicultural operation. The lack of response in many instances, however, can be traced to the prior condition of the trees, the experimental techniques, or more frequently, to the analysis and interpretation of the results.
Inheritance must also be taken into consideration, and it must constantly be born in mind that all trees are predisposed to assume a certain form. Environmental influences and cultural practices can only modify the basic tree form predestined by heredity.
Points of inflection along the stem present a major problem in specifying stem form. Besides the two major points, one in the butt region and the other somewhere in or close to the crown base, there are many minor points of inflection along the bole. In other words, tree form is complex. This makes it difficult to specify the profile of a single stem, let alone to specify a profile to represent all stems. Nevertheless, researchers have attempted to do this through two broad approaches:
Support for both the Nutritional and Water Conducting theories has waned in recent decades.
Metzger proposed that the stem was a beam of uniform resistance to bending, anchored at the base and functioning as a lever arm. A horizontal force will exert a strain on the beam that increases toward the point of anchorage, and if the beam is composed of homogeneous material the most economical shape would be a beam of uniform taper. Metzger demonstrated that this taper approximates the dimensions of a truncated cubic paraboloid (height against diameter^3 is linear). He confirmed his theory for many stems, particularly conifers.
Gray (1956), an Australian forester, questioned Metzger's assumption that the tree stem is anchored firmly to the ground, and contended that stem form conforms more closely to the dimensions of a quadratic paraboloid (height against diameter^2 is linear). A stem of this shape would be consistent with the mechanical requirements of a tree, not only to horizontal wind pressure but to other forces acting on the stem.
There have been many attempts to prove and disprove the theory (e.g. Larson, B. 1963. For. Sci. Monogr. No. 5). Generally, it is accepted that the theory provides a realistic interpretation of tree form with respect to strength requirements and support function. Though the theory has survived experimental study reasonably well, it does not provide an adequate physiological explanation of the observed facts.
It provides a physiological explanation of how a tree grows and why trees differ in the way they do, but it does not specify the particular shapes trees may have under varying circumstances.
The hormonal theory offers the most promising approach to the stem form problem. It provides a physiological basis for the nutritional as well as the functional theories of stem form.
Most commonly, the form factor of trees is based on a cylinder. Thus, the product of tree basal area, tree height and cylindrical form factor should give tree volume.
The stimulus for the determination of form factors was the recognition of the strong resemblance of a tree stem to standard geometric solids. The hope was that ratios would be established which could be used to convert easily computed volumes of standard solids into tree volumes.
Several types of form factor exist:
As with form factor there are several types of form quotient, the most common being absolute form quotient which is the ratio of the diameter at half height above breast height to dbh. This is commonly written as d5/d0 and, conventionally, is expressed as a decimal, e.g. 0.70. Form quotient values are grouped into form classes, e.g. Form Class 70 comprises trees with F.Q.s from 0.675 to 0.724.
Absolute form quotient (and classes):
Jonson used this formula to compile tables giving, for various form classes, tree diameters at percentile heights as percent of dbh. Thus, the profile of a stem is defined and so, automatically, is its form factor.
Following Jonson, other workers (Wright, Behre, etc.) investigated other stem profile equations but none was found to be entirely satisfactory.
Gray developed his hypothesis of stem form based on empirical evidence (from numerous sample trees covering a wide range of species) that over a large part of a coniferous stem, height above ground is directly proportional to the square of diameter at that point. The practical application of his hypothesis is the taper line method of volume estimation.
Most are of these equations empirical and they do not cater adequately for the variability found in any one population. The nature of the stem profile is compound and it would seem that it can only be described adequately by separate expressions for the top section, main bole and buttswell region. In addition, the sections of stem described by these separate expressions vary with species, age, size, stand condition and environment.

Grosenbaugh (1966), following a comprehensive study of stem form, writes (Forest. Chronicle 42: 444-57):
Many mensurationists have sought to discover a single, simple two-variable function involving only a few parameters which could be used to specify the entire tree profile. Unfortunately, trees seem capable of assuming an infinite variety of shapes, and polynomials (or quotients of polynomials) with degree at least two greater than the observed number of inflections are needed to specify variously inflected forms.Thus, it appears unwise to derive complicated relationships to characterise the tree profile. This notion is corroborated by several researchers who, after studying various multivariate analysis techniques, concluded that the use of simple functions, sorting, and graphical methods is adequate for many uses in operations and research.Furthermore, coefficients would vary from tree to tree in ways that could only be known after each tree has been completely measured. Thus, explicit analytic definition (of the tree profile) requires considerable computational effort, yet lacks generality......
Each tree must be regarded as an individual that must be completely measured, or else as a member of a definite population whose average form (profile) can only be estimated by complete measurement of other members of the population selected according to a valid sampling plan.... Hence, polynomial analysis may rationalise observed variation (in the stem profile) after measurement, but it does not promise more efficient estimation procedures."
A majority of coniferous stems in Australia have a FQ of about 0.7 so there is considerable evidence for the generality of Gray's hypothesis. The hypothesis also holds well for most, but not all, native broadleaf species from above the buttswell region to crown break.
![[Index]](index1.GIF)
![[Help]](help1.GIF)
![[Authors]](author1.GIF)
http://online.anu.edu.au/Forestry/mensuration/SHAPE.HTM
Cris.Brack@anu.edu.au
Tue, 7 Jan. 1997